オートファゴソーム形成の分子機構の形態学的解析(平田恵理・鈴木邦律)
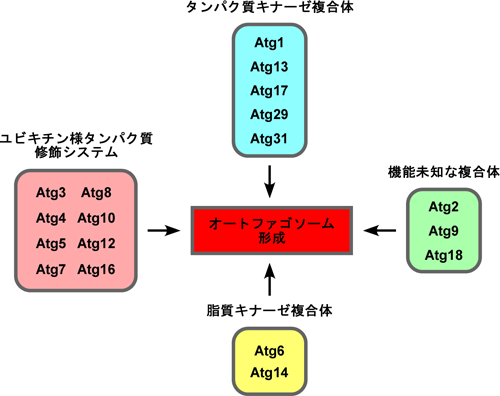
細胞が栄養飢餓を感知すると、隔離膜と呼ばれる小さな袋状の膜が細胞質に出現する。隔離膜は湾曲しつつ伸展し、やがて伸展した隔離膜の末端が閉じることによって、被分解物を内包したオートファゴソームが完成する。現在のところ、オートファゴソーム形成に関わる18種類のATG(autophagy-related)遺伝子が知られており、それらは少なくとも4つの機能群を構成して機能していると考えられている(図1)。最近になって、これらのAtgタンパク質はどれも隔離膜形成に必要なことが分かってきた。私たちの研究室では、出芽酵母において隔離膜を可視化する手法を開発し、各々のAtgタンパク質が隔離膜形成中に細胞内局在を変化させる様子をライブイメージング(生細胞観察)により解析し、隔離膜を経てオートファゴソームが形成されていく分子機構の全容を解明すべく研究を進めている。
 |
|
図1.オートファゴソーム形成に関わる18種類のAtgタンパク質の機能群 |
- Fine mapping of autophagy-related proteins during autophagosome formation in Saccharomyces cerevisiae.
- Kuninori Suzuki, Manami Akioka, Chika Kondo-Kakuta, Hayashi Yamamoto and Yoshinori Ohsumi.
- J. Cell Sci.
- 2013 Jun 1;126(11):2534-2544. Epub 2013 Apr 2.
- Autophagosome formation can be achieved in the absence of Atg18 by expressing engineered PAS-targeted Atg2.
- Takafumi Kobayashi, Kuninori Suzuki and Yoshinori Ohsumi.
- FEBS Lett.
- 2012 Jul 30;586(16):2473-8. Epub 2012 Jun 19.
- Current knowledge of the pre-autophagosomal structure (PAS).
- Kuninori Suzuki and Yoshinori Ohsumi.
- FEBS Lett.
- 2010 Apr 2;584(7):1280-6. Epub 2010 Feb 5.
- Molecular machinery of autophagosome formation in yeast, Saccharomyces cerevisiae.
- Kuninori Suzuki and Yoshinori Ohsumi.
- FEBS Lett.
- FEBS Lett. 2007 May 22;581(11):2156-61. Epub 2007 Mar 12.
- Characterization of the Atg17-Atg29-Atg31 complex specifically required for starvation-induced autophagy in Saccharomyces cerevisiae.
- Yukiko Kabeya, Nobuo N. Noda, Yuko Fujioka, Kuninori Suzuki, Fuyuhiko Inagaki and Yoshinori Ohsumi.
- Biochem. Biophys. Res. Commun.
- 2009 Nov 27;389(4):612-5. Epub 2009 Sep 13.
- Hierarchy of Atg proteins in pre-autophagosomal structure organization.
- Kuninori Suzuki, Yuka Kubota, Takayuki Sekito and Yoshinori Ohsumi.
- Genes Cells
- 2007 Feb;12(2):209-18.
- The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation.
- Kuninori Suzuki, Takayoshi Kirisako, Yoshiaki Kamada, Noboru Mizushima, Takeshi Noda and Yoshinori Ohsumi.
- EMBO J.
- 2001 Nov 1;20(21):5971-81.
関連論文
オートファゴソーム内容物の網羅的解析(王・鈴木邦律)
オートファジー不能の表現型はAPの積荷が分解できないことに起因する。すなわちオートファジーの生理的意義を理解するにはオートファジーによって何が分解されているのか、言い換えるとAPの内容物が何であるのかを解明する必要がある。我々はAPの選択的積荷およびその分子機構を次々と明らかにしてきたが、未だオートファジーの生理的意義の全容解明にはほど遠い。現在生化学的に単離したAPの内容物を網羅的に同定するプロジェクトを進め、オートファジーによる被分解物の同定を通じてその生理的意義に迫ろうとしている。
- Proteomic profiling of autophagosome cargo in Saccharomyces cerevisiae.
- Suzuki K, Nakamura S, Morimoto M, Fujii K, Noda NN, Inagaki F, Ohsumi Y.
- PLoS One.
- 2014 Mar 13;9(3):e91651.
- Selective autophagy in budding yeast.
- Kuninori Suzuki.
- Cell Death Differ.
- 2013 Jan;20(1):43-8. Epub 2012 Jun 15.
- Selective autophagy regulates insertional mutagenesis by the Ty1 retrotransposon in Saccharomyces cerevisiae.
- Kuninori Suzuki, Mayumi Morimoto, Chika Kondo and Yoshinori Ohsumi.
- Dev. Cell
- 2011 Aug 16;21(2):358-65.
- Selective transport of α-mannosidase by autophagic pathways: identification of a novel receptor, Atg34p.
- Kuninori Suzuki, Chika Kondo, Mayumi Morimoto and Yoshinori Ohsumi.
- J. Biol. Chem.
- 2010 Sep 24;285(39):30019-25. Epub 2010 Jul 16.
- Selective transport of α-mannosidase by autophagic pathways: structural basis for cargo recognition by Atg19 and Atg34.
- Yasunori Watanabe, Nobuo N. Noda, Hiroyuki Kumeta, Kuninori Suzuki, Yoshinori Ohsumi and Fuyuhiko Inagaki.
- J. Biol. Chem.
- 2010 Sep 24;285(39):30026-33. Epub 2010 Jul 21.
- Studies of cargo delivery to the vacuole mediated by autophagosomes in Saccharomyces cerevisiae.
- Kuninori Suzuki, Yoshiaki Kamada and Yoshinori Ohsumi.
- Dev. Cell
- 2002 Dec;3(6):815-24.
関連論文
オートファジックボディ分解機構の解析(白井・鈴木邦律)
APは細胞内分解コンパートメントである液胞と融合した後、内膜からなる小胞として液胞内腔に放出される。この小胞はオートファジックボディ(AB)と呼ばれ、野生株ではABの寿命は1分に満たず、液胞内でたちまち崩壊する。ABの崩壊には液胞内のタンパク質分解酵素であるPep4pやPrb1pが必要であるが、AB表面にはタンパク質がほとんど存在しないことから、AB膜を直接分解する酵素の寄与が考えられている。Atg15はリパーゼ様の酵素であり、Atg15を欠いた変異株ではABが液胞内に蓄積することから、Atg15がABを直接分解している可能性が示唆されている。我々はAtg15によるAB分解のメカニズムを解明すべく研究を進めている。
- Studies of cargo delivery to the vacuole mediated by autophagosomes in Saccharomyces cerevisiae.
- Kuninori Suzuki, Yoshiaki Kamada and Yoshinori Ohsumi.
- Dev. Cell
- 2002 Dec;3(6):815-24.
関連論文
出芽酵母の種々のオルガネラの形態を数値化し解析する(鈴木邦律)
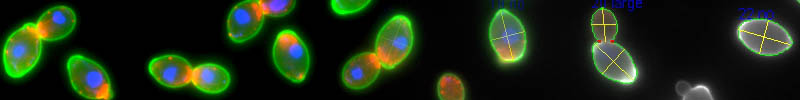
出芽酵母は単細胞ではあるが、真核細胞のモデル生物として広く使用されている。それは、核・小胞体・ミトコンドリア・液胞(リソソーム)・ペルオキシソーム・脂肪滴などのオルガネラが高等真核細胞と同様なメカニズムで形成・維持されているからに他ならない。加えて、出芽酵母は遺伝子破壊や遺伝子導入が容易であり、生育に必要でない遺伝子を一通り破壊した株を容易に入手可能である。当研究室では核・アクチン・細胞壁を可視化した出芽酵母の形態情報をCalMorphソフトウェアによって数値化することで、遺伝子の機能と形態変化との関係を探ってきた。この手法を上記のオルガネラに拡張し、超高次元形態情報に基づいた表現型解析を可能にすべく解析手法の開発を進めている。
- Organelle acidification is important for localisation of vacuolar proteins in Saccharomyces cerevisiae.
- Risa Matsumoto, Kuninori Suzuki and Yoshikazu Ohya.
- Protoplasma
- 2013 May 25. [Epub ahead of print].
関連論文
オートファジーの全過程を試験管内で再構成する(鈴木邦律)
オートファジーをAPの一生と捉えると、(1)AP形成、(2)APの液胞との融合とABの出現、(3)ABの崩壊のステップに分割して考えることができる。当研究室では、それぞれのステップを試験管内で再構成し、オートファジーの全過程を試験管内で再現することを目指し、研究を進めている。